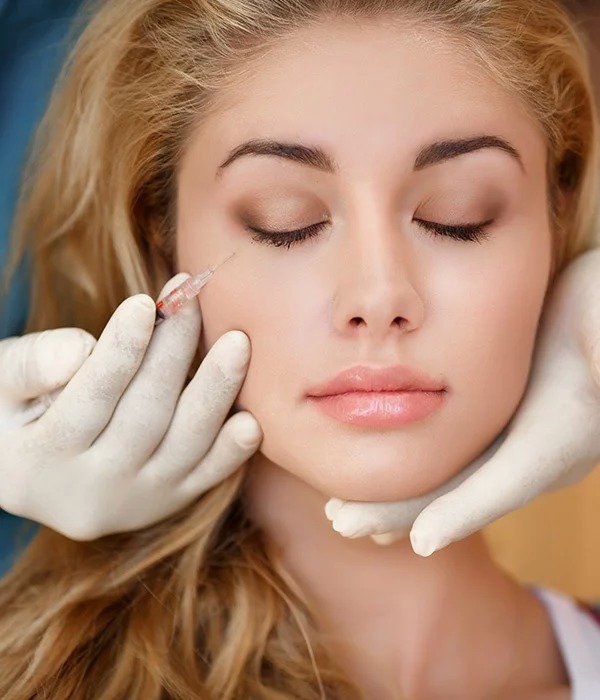
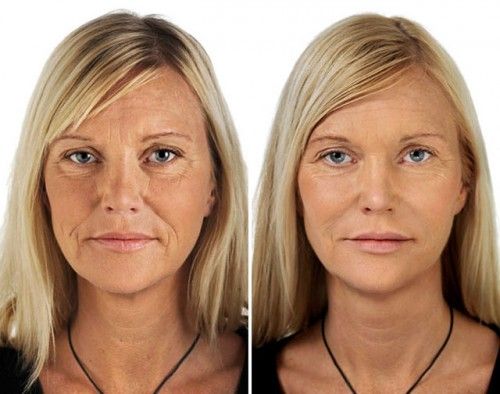
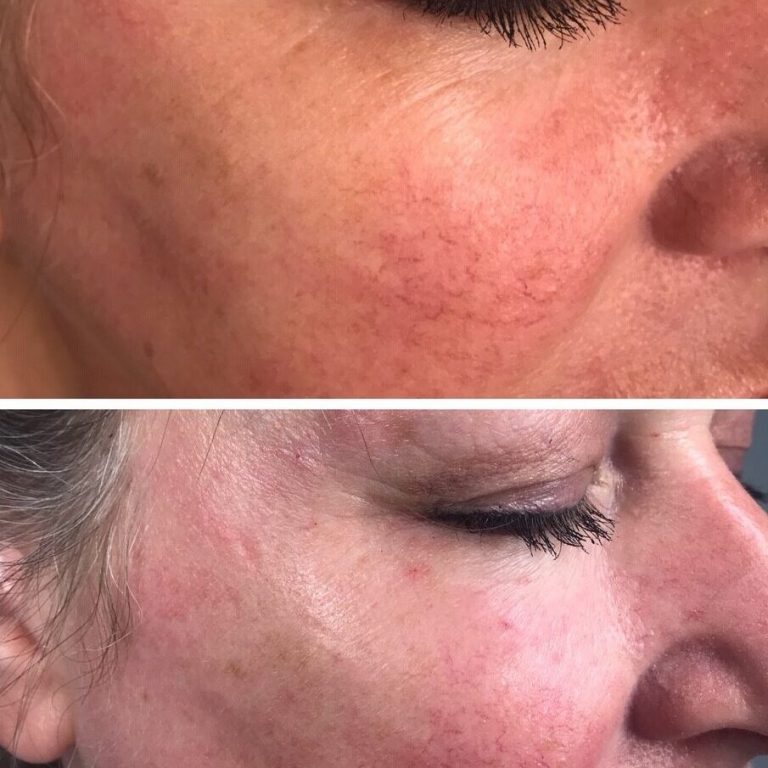
The eyes and the area around them are the first thing you notice when determining the age of your companion. So let’s find out how you can lose a few years of age by rejuvenating that periorbital area around your eyes. You can get more useful information about fillers at https://majormedicalsolutions.com/.